Hva er lysveileder.no?
På lysveilder.no finner du alle Lyskultur sine veiledere innenfor fagfeltet lys og belysning. Veilederne tar for seg mange ulike tema og de ulike standardene som benyttes for faget. Nedenfor ser du hvilke publikasjoner som er tilgjengelig for abonnenter.
Gjennom et abonnement har du tilgang på alt innhold, men det er også mulig å bestille enkeltstående publikasjoner fra Standard Norge.
I tillegg har Lyskultur laget fagark med spesifikke tema, samt noen mer omfattende hefter og utgivelser. Disse er kun tilgjengelige på lysveileder.no.
Flere byggeprosjekter angir i beskrivelser, under "Generelle krav" at en eller flere av våre veiledere skal følges, så et abonnement er nyttig også for byggeiere og entreprenører foruten rådgivere og leverandører.
Som medlem av Lyskultur får man rabatt på abonnement på lysveileder.no.
Gjeldende priser:
Ikke medlem:
| Per bruker: | 5 700,- |
Medlem i Lyskultur:
| Personlig abonnement (en bruker): | 1 990,- |
| 2-9 ansatte: | 4 300,- |
| 10-49 ansatte: | 8 550,- |
| 50-99 ansatte: | 18 350,- |
| 100+ ansatte: | 21 800,- |
| Foreninger, non-profit, kommuner og skoler (en bruker): | 1 990,- |
Abonnementet løper og fornyes automatisk til gjeldende pris inntil det blir sagt opp. Avbestilling må skje skriftlig, og være Lyskulturs administrasjon i hende før 31.10 for utmelding påfølgende år.
Lysboken og luxtabeller
Grunnleggende begreper og kriterier
1A
1A Lysboken
Dette er Lyskulturs mest omfattende publikasjon, og bør være av interesse og nytte for enhver som studerer eller jobber med lysfaget.

1B
1B Luxtabell
Veilederen har som formål å gi råd og veiledning for riktig bruk av belysning i ulike arbeidssituasjoner og for ulike funksjoner.

1C
1C Luxtabell
Publikasjonen gir retningslinjer for aktuelle krav til belysningen av utendørs arbeidsplasser, men også for annen virksomhet tilknyttet utendørs aktiviteter.

Lysveiledere
Veiledere for ulike belysningsstandarder og for planlegging av belysning
3
Idrettsbelysning
Publikasjonen erstatter Lyskulturs publikasjon Nr. 3 – Idrettsbelysning fra 2013.

7
Nødlys- og ledesystemer
Formålet med publikasjonen «Nødlys/ledesystemer» er å være en guide i prosessen rundt søknad, prosjektering, utførelse og bruk samt kontroll og vedlikehold av landbaserte systemer.

10
Marine lighting
Norge er verdensledende på marin belysning. Skipsbyggingsnæringen er solid, og belysning følger med som en naturlig del av dette. For første gang på 40 år kommer nå publikasjonen 10 – Marine Lighting i ny og revidert utgave.

11
Belysning for eldre og svaksynte
Med en økende andel eldre i samfunnet er det viktig at både offentlige og private rom og steder er utformet slik at det trygt og sikkert kan brukes av alle, fra ung til gammel.

12
Kontorbelysning
Formålet med publikasjonen Kontorbelysning er å gi råd og veiledning for hvordan planlegging og prosjektering av gode belysningsanlegg på arbeidsplasser kan gjennomføres. Den gir videre nyttige av prosesser frem mot vellykkede løsninger.

17
Belysning i museer, gallerier og utstillinger
Formålet med veilederen Belysning i museer, gallerier og utstillinger er å gi råd og veiledning for hvordan planlegging og prosjektering av gode belysningsanlegg i museer, gallerier og utstillinger kan gjennomføres. Veilederen skal gi inspirasjon og ideer til funksjonelle belysningsløsninger som også tar hensyn til energieffektivitet og bærekraft.

20
Lys i læringsmiljø
Publikasjonen Lys i læringsmiljø er ment å være et verktøy for utdanningssektoren i arbeidet med å skape gode skoleanlegg.

21
Dagslys i bygninger
Dagslys er jordens essensielle grunnleggende lyskilde, gir verdifulle tilskudd til trivsel og innemiljø.

24
Lysstyring
Formålet med publikasjonen er å gi råd, veiledning og kompetanse til alle som prosjekter, leverer, installerer eller bestiller lysstyringsanlegg.

25
Belysning av veier, gater og byrom
Veilederen gir god veiledning i de ulike faser ved planlegging, installasjon og drift av belysningsanlegg for vei, gate og byrom.

26
Belysning og universell utforming
Lyskulturs publikasjon nr. 26 Belysning og universell utforming er en helt ny publikasjon som skal bidra til felles forståelse for hvordan hensynet til universell utforming kan ivaretas som en del av en samlet belysningsløsning.

Faktaark
Korte dokumenter om spesifikke tema
F01
F01 Vedlikeholdsfaktor*(MF)
Vedlikeholdsfaktoren (Maintenance Factor) er et kjent begrep for de som planlegger belysningsanlegg. Denne faktoren skal benyttes som en reduksjonsfaktor, og sikrer at den planlagte belysningsinstallasjonen til enhver tid skal gi et tilstrekkelig lysnivå for den bruken anlegget er tiltenkt.
F2
LED og nødvendig dokumentasjon
Dette faktaarket er ment å gi aktørene i det profesjonelle belysningsmarkedet noen retningslinjer for hvilken type dokumentasjon man trenger for å sammenlikne leverandører av produkter med LED.
F3
Dagslysfaktor
Dette faktaarket er laget for å forklare begrepet dagslysfaktor, kravene til denne, samt hvordan dagslysfaktor kan beregnes og måles. Faktaarket omhandler ikke de kvalitative forholdene med hensyn til dagslys i bygninger, systemer for solavskjerming eller dagslystransport.
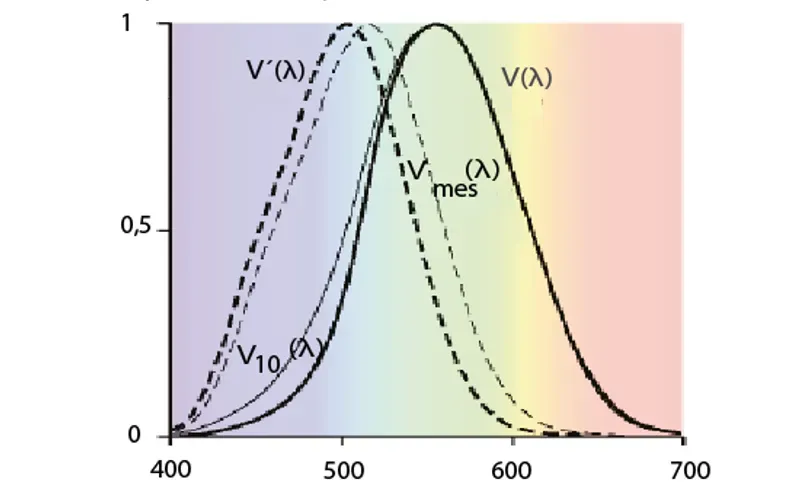
F4
Mesopisk fotometri – skumringslys
Overgangen til å benytte metallhalogen og LED til utendørs belysning er ledsaget både av seriøse studier og av mer lettvinte spekulasjoner om den visuelle nytten av å benytte hvitt lys i stedet for gult lys. To rapporter fra den internasjonale belysningskommisjonen, CIE 191:2010 og CIE 206:2014 gir oss et foreløpig faktagrunnlag, som er oppsummert i dette faktaarket.
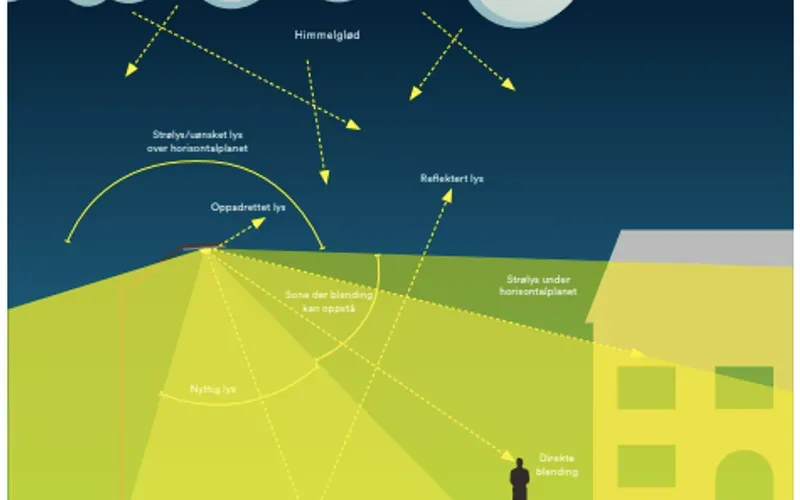
F6
Lysforurensning
Lysforurensing er den totale summen av uønskede og uheldige effekter av kunstig lys. Enkelt kan det betegnes som lite tilpasset eller overflødig elektrisk belysning. Det har en rekke negative konsekvenser, som forringede muligheter for observasjon av nattehimmelen, forstyrrelse av økosystemer og ikke minst energisløsing. Heldigvis er dette en type forurensning som det er relativt lett å gjøre noe med, gjennom optimalisering av lysanleggene.
F7
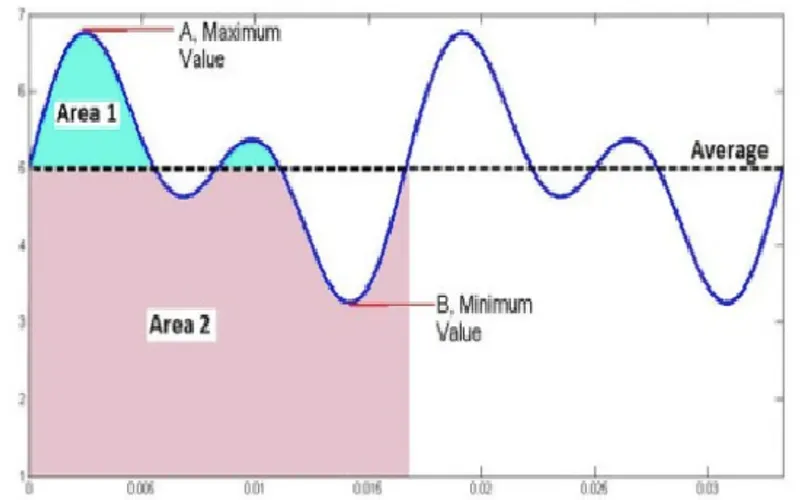
Led og flimmer
Den internasjonale belysningskommisjonen (CIE) arbeider for tiden med et nytt konsept for karakterisering og måling av tidsmodulerte belysningssystemer, herunder flimmer ifm. LED-belysning. Dette arbeidet ligger imidlertid et par år frem i tid, og dette faktaarket er derfor ment som hjelp/referanse inntil nye beregningsmetoder og retningslinjer er fastlagt av CIE. Faktaarkets status er av denne grunn kun å betrakte som midlertidig.
F8
Informasjon om LED-belysning
Hvorfor dette informasjonsheftet om LED-belysning? Dette er en oppdatering av Informasjonsheftet som ble publisert på www.efo.no i 2009. Dette faktaark er utarbeidet av EFO i 2014 og oppdateringen omfatter de fleste kapitlene. Mens utgaven fra 2009 var rettet til installatører er denne utgaven også tiltenkt andre brukere av LED eller alternativ teknologi.

F9
BREEAM-krav for lys og belysning
BREEAM (Building Establishment’s Environmental Assessment Method) er en internasjonal anerkjent metode for å måle et byggs bærekraftige egenskaper. Formålet er å motivere til bærekraftig planlegging og bygging gjennom hele byggeprosjektet, fra tidligfase til overlevert bygg.
F10
Kvalitetssikring av belysningsanlegg
Sjekkliste for belysningsanlegg. Hvordan gjøre en etterkontroll for å påse at det som blir levert er i henhold til det som er bestilt? For å kontrollere leveransen i forhold til oppgaven, er det laget en enkel sjekkliste. Sjekklisten tar for seg tre ulike stadier i prosjektforløpet, i tillegg til grunnlaget for prosjekteringen.
F11
Veileder i sikringsbelysning
Denne veilederen gir en innføring i hvordan man tar hensyn til sikkerhet ved planlegging og etablering av belysning.
Den retter seg mot brukere, tiltakshavere, virksomhetseiere og prosjektledere som ikke har sikringserfaring. Vi ønsker med dette å skape bevissthet rundt emnet og begrepet sikringsbelysning og vise behovet for bruk av faglig kompetanse. Dessuten gis det eksempler på tiltak og løsninger.
F12
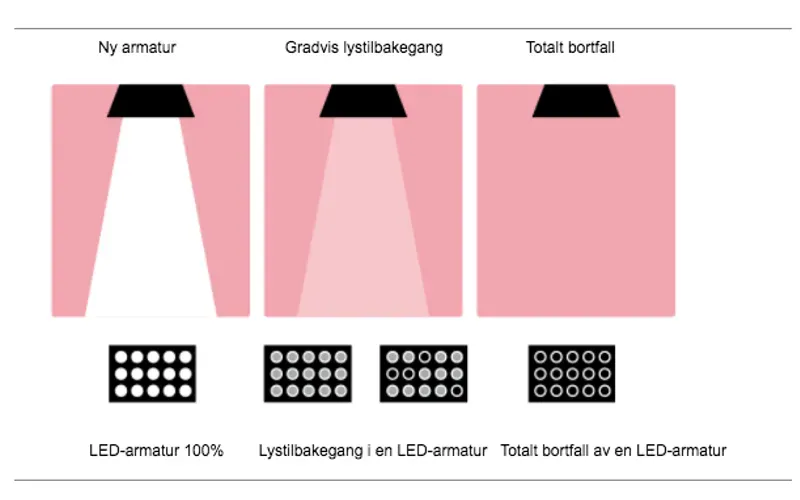
LED-armaturer for innendørs bruk
For å kunne sammenligne ulike LED-armaturer og lysberegninger på en korrekt og rettferdig måte er det viktig at relevante data oppgis og blir sammenlignet på likt grunnlag, både mekaniske egenskaper og lystekniske data.
F13
LED-armaturer for utendørs bruk
For å kunne sammenligne ulike LED-armaturer og lysberegninger på en korrekt og rettferdig måte er det viktig at relevante data oppgis og blir sammenlignet på likt grunnlag, både mekaniske egenskaper og lystekniske data.
Øvrige utgivelser
Lysrelaterte utgivelser fra Lyskultur og ulike samarbeidspartnere
Godt lys i boligen
Lys er avgjørende for mange gjøremål i boligen. Lys er viktig for sikkerheten. Lys er med på å skape en sammenhengende helhet i hjemmet, og har betydning for stemning, trivsel og atmosfære. Kort sagt: Lys skaper liv.

Lys på stedet
God utendørsbelysning gjør byene og tettstedene våre triveligere, tryggere og mer tilgjengelige. Dessuten gir riktig lysbruk også mulighet til å spare penger.

Bedre belysning – Prosessveileder
Denne prosessveilederen er utarbeidet av Lyskulturs faglige komité (NLK) som har ansvar for utarbeidelse av Lyskulturs faglige publikasjoner og standardiseringsarbeid. Lyskultur er Norges ledende kompetansenettverk innen lys og belysning.
